トップ>研究内容>研究成果(long version)>3. 樹状突起-分岐の複雑度や受容野のサイズを調節する分子機構-
3. 樹状突起-分岐の複雑度や受容野のサイズを調節する分子機構-
研究の背景と成果
神経細胞は高度に極性化された細胞であり、2種類の突起(樹状突起と軸索)を伸展させる。 樹状突起はシナプス入力または感覚入力を受容するアンテナとして働き、一般的に軸索よりも短くはるかに複雑に枝分かれする。ニューロンはクラスごとに特徴 的な樹状突起パターンを発達させるが、その驚くばかりの多様性がどのように調節されているかは未解決の問題である。軸索はニューロンのクラス間で形態上の 差異が小さいため、多数の軸索を集団として扱う実験系を用いて研究が発展してきた。クラス毎に形状が大きく異なり、狭い空間内で複雑に分岐する樹状突起の パターン形成を研究するには、一細胞の解像度で、しかも再現性よく同じクラスのニューロンを可視化する必要がある。そこで我々は、以下に述べるように、 ショウジョウバエの dendritic arborization (da) neuron をモデルとして、個体を生かしたまま樹状突起を一細胞レベルで観察できる系を作り上げた。
主要な論文1
- Kaoru Sugimura(杉村薫), Daisuke Satoh, Patricia Estes, Stephen Crews, and Tadashi Uemura. Development of morphological diversity of dendrites in Drosophila by the BTB-zinc finger protein Abrupt. Neuron, 43: 809-822 (2004).
da neuron は、分岐の複雑度と樹状突起がおおう領域(受容野)のサイズの順に応じてクラスI?IVに分類されており、 樹状突起形態の多様性を解析 する上で優れたモデル系を提供する (Sugimura et al. 2003;図5と図6)。クラスI ニューロンとは対照的に、クラスIV ニューロンは幼虫期を通してより高次の分岐を展開し続ける。また、レーザーを用いて突起を切断すると、同じクラスIVニューロンの他の枝や、隣接するクラ スIVニューロンの枝が、切断された突起が占めていた受容野を素早く埋める応答を示す。
各クラスに特徴的な 突起形態を発達させる仕組みを明らかにする目的で、特定の クラスに選択的に発現する遺伝子を探索した。その結果、 Zinc finger を持つ転写調節因子Abrupt(Ab) が、クラスI ニューロンに特異的に発現していることを見出した(神経前駆細胞での発現レベルは低く検出は困難だった)。他のクラスに属するニューロンにおいて、その ニューロンが誕生した後に Ab を強制発現させると、その細胞はクラスIのように分岐の複雑度が低く、かつ受容野の狭い樹状突起を形成させた。クラスIVニューロンに発現させた場合に は、レーザーによる突起切断に対する応答も失われた。対照的に、 ab 機能喪失変異により、クラスI ニューロンは正常よりも複雑に分岐する突起を形成した。以上の結果から Ab は、(神経前駆細胞のレベルでクラス特異性を決定するのではなく、ニューロンが生まれた後で)クラスI に特徴的な樹状突起パターンを直接制御する可能性が支持された (Sugimura et al. 2004)。
Ab とは対照的に、クラスIVニューロンで選択的に発現する転写調節因子 Knot (Kn) を見出した。Kn をクラスI ニューロンで強制的に発現させると、その樹状突起は正常細胞に比べはるかに複雑に分岐した。よってKn の発現はクラスIV に特徴的な突起パターンの形成を賦与するらしい(未発表データ)。以上の結果から、転写調節因子群によるクラス選択的な突起パターンの形成機構が存在する ことが示された(図7)。Ab や Kn に対して、ホメオドメインを持つ Cut (Ct) はクラスIIIでの発現が最も高く、そのクラスに特徴的な突起形態の獲得に必要かつ十分であることが他のグループにより示されていた。Ct や Kn についてはほ乳類のホモログが報告されており、我々は Ab のほ乳類ホモログの候補を探索している。それらの転写調節因子群が、ほ乳類神経系においても 樹状突起形態の多様性を調節するのか興味が持たれる。
関連論文
- Kaoru Sugimura, Misato Yamamoto, Ryusuke Niwa, Daisuke H. Sato, Satoshi Goto, Misako Taniguchi, Shigeo Hayashi, and Tadashi Uemura. Distinct developmental modes and lesion-induced reactions of dendrites of two classes of Drosophila sensory neurons. J. Neuroscience, 23:3752-3760 (2003).
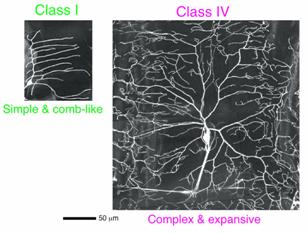
図6 クラスIとクラスIVに属するda neuronの例
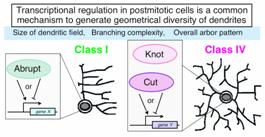
図7 転写調節因子群による、クラス選択的な樹状突起パターン形成の概念図
主要な論文2
-
Misato Yamamoto(山本美暁), Ryu Ueda, Kuniaki Takahashi, Kaoru Saigo, and
Tadashi Uemura. Neuron, glia, and epidermal cells: tricellular
regulation of neuronal morphology through an immunoglobulin superfamily
molecule Neuroglian. Current Biology, 16:1678-83 (2006).
樹状突起のパターン形成を含めて、神経細胞の形態形成における細胞間認識の役割を解明することを狙い、 da neuron で発現している細胞膜タンパク質を探索した。その結果、Ig super family の中でも L1 family に属する Neuroglian (Nrg) に注目した。Nrg は胚期や終齢幼虫期においてda neuronの樹状突起や軸索だけでなく、表皮細胞と da neuron の軸索束を包むグリア細胞にも発現していた。突然変異体の解析などから、da neuron とグリア、あるいは da neuron と表皮細胞境界とのNrg依存的な相互作用が、軸索分岐抑制と樹状突起パターン形成のそれぞれに重要であることが示された。
他の研究実施内容:突然変異体のスクリーニングなど
上記のアプローチに加えて、突起の分岐や伸長の異常を指標とする突然変異体のスクリーニングを行っている。スク リーニング法の一つとして、 da neuron で GFP を発現するトランスジェニック系統のゲノムに突然変異を誘発し、突起パターンに異常を示す変異体を分離した。合計2170 系統の致死変異株を分離し、各々について樹状突起を観察した。その結果少なくとも 17 系統において、高次の枝の形成が抑えられる異常や突起伸長方向の異常など、様々な表現型が検出できた。責任遺伝子のクローニングを進めている。正常なクラ スIVニューロンの樹状突起は、細胞体から遠い領域においてさかんに分岐するが、ある突然変異を持つニューロンの突起は、逆に近い領域で多数の側枝を生や すことがわかった。この突然変異の表現型を指標にすることで、生体内で働く分岐装置の分子的実体を明らかにできると思われる。
突然変異体のスクリーニングに加えて、 樹状突起のパターン形成を調節する可能性のある da neuron 内の現象や、da neuron と他の細胞間とのコミュニケーションを解析するために、種々の蛍光タンパク質マーカーを発現するトランスジェニックフライの作製を試みている。例えば、 樹状突起形成において細胞膜成分がどのように供給されているのか、そして種々のオルガネラはどのような役割を果たすのかは、生体内ではほとんど解析されて いない。そこでゴルジ体などを可視化できるトランスジェニックマーカーを作製しつつあり、その新たなマーカーを利用したイメージングを試みている。また、 da neuron とその周囲に存在する血球系細胞群との相互作用にも着目し解析を始めている。
今後の展開
樹状突起形態の多様性を解明する目的で、転写調節因子 Ab あるいは Knot の標的遺伝子の分離を目指した実験を開始している。マイクロアレイ解析に十分な量の、特定クラスに属するニューロンを単離することが技術的に困難であるこ とが予想されたので、胚あるいは幼虫全体で解析が可能か予備的な実験を行っている。
標的遺伝子を直接探索する試みを続ける一方で、標的遺伝子は複数あることが予想されるので、クラス毎に選択的に用いられる仕組みと、複数のクラスに共通に 用いられる機構との接点を探り、遺伝子プログラム全体の解明を目指す必要がある。また、クラス IV ニューロンについては、その振る舞いを説明できる数理モデルを検討しており、実験の現場へのフィードバックを目指している。
樹状突起パターンの多様性はニューロンのクラスごとに特有な機能を支えており、神経系が様々な情報を受容し処理す るために不可欠だと考えられている。それぞれの神経回路の中で、特徴的な突起パターンを発達させたニューロンが適材適所に配置されなければ、ヒトの脳は思 考や感情、運動、知覚などの情報を適切に処理しきれないであろう。この突起パターンの多様性を支える遺伝子プログラムの全体像に迫るには、様々なアプロー チを用いて da neuron の利点を生かし続ける必要がある。本研究は細胞レベルでの視点からスタートしているが、da neuron の機能を個体の行動をアウトプットとして調べるアプローチも検討している。da neuron を用いた今後の研究から、(個々の遺伝子の発見だけではなく)突起パターン形成プログラムの一部分でも明らかになれば、哺乳類ニューロンの神経回路形成へ の研究に大きな波及効果を生み出せるであろう。